Giving Voice to Stomata Differentiation
Torii K. U. (2007)
Cell 128(3): 419 – https://doi.org/10.1016/j.cell.2007.01.025 –
The generation of asymmetry is a key event in many developmental processes, including cell differentiation, embryonic axis specification, and organogenesis. Recent research highlights the identification of factors needed to establish stomata during plant development and the role that timing plays in directing left-right asymmetry during nervous system development in zebrafish. Other recent studies elucidate unexpected connections between two kinds of asymmetry, cellular and organismal, and how asymmetry may be related to the regulation of chromatid segregation and DNA repair.
Giving Voice to Stomata Differentiation
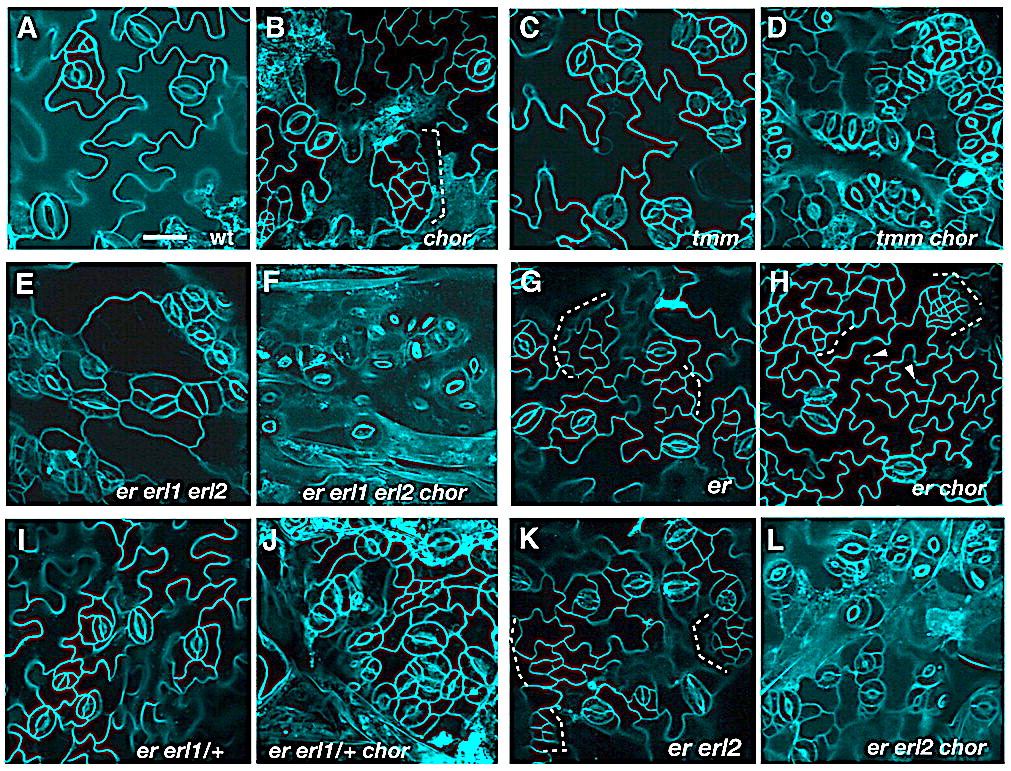
Stomata are the pores that mediate gas and water exchange in plants. They are formed by a symmetrical pair of guard cells that regulate the opening and closing of the pore, yet the specification of the guard cell precursor relies upon a series of asymmetric cell divisions beginning in a single meristemoid mother cell. Two groups (MacAlister et al., 2006; Pillitteri et al., 2006) now report that three related basic helix-loop-helix transcription factors (SPEECHLESS, MUTE, and FAMA) act sequentially in the lineage that gives rise to guard cells in Arabidopsis thaliana. As their names imply, loss of either SPEECHLESS or MUTE leads to seedlings that develop leaves devoid of stomata. In the first step of guard cell specification, SPEECHLESS facilitates the first asymmetric division of the meristemoid mother cell to establish the meristemoid. MUTE then terminates the subsequent rounds of asymmetric cell division to promote the differentiation of the meristemoid into a guard mother cell. The third factor FAMA, identified in a previous study, promotes the final transition from guard mother cell to guard cells. Future work may identify the targets regulated by these transcription factors. Of particular interest would be uncovering the genes regulated by SPEECHLESS that drive the asymmetric division of the meristemoid mother cell. The findings of MacAlister et al. and Pillitteri et al. also reveal a striking parallel between the differentiation of stomata and developmental processes in animals, including myogenesis and neurogenesis, which also require the sequential activity of related basic helix-loop-helix transcription factors.
C.A. MacAlister et al. (2006). Nature. Published online December 20, 2006. 10.1038/nature05491.
L.J. Pillitteri et al. (2006). Nature. Published online December 20, 2006. 10.1038/nature05467.










{kind=link}
{kind=link}
You must be logged in to post a comment.