The science of the stomata of plants: a continuously growing list of references, abstracts and illustrations, helping researchers to data on publications.
Some studies have shown that stomata in detached epidermis behave differently, both quantitatively and qualitatively, from those in the intact leaf. Stomata in the intact leaf are very sensitive to environmental factors such as light, CO2 and osmotic stress, but stomata in detached epidermis are less sensitive to these factors than those in the intact leaf. Stomata in the intact leaf open in the light and close in the dark, whereas previously opened stomata in detached epidermis do not close on transfer to darkness and they are dependent on the KCl concentration in the incubation medium.
Electrophysiological results also demonstrate different responses of guard cells in the intact leaf and detached epidermis. The electrical potential across the membrane of the guard cell in the intact leaf was hyperpolarised in response to light and CO2. However, there were no changes in the membrane potential difference in response to light and CO2 in detached epidermis. In CCCP-infiltrated leaf discs, the guard cell membrane was conversely depolarised by light, indicating that the guard cell membrane hyperpolarisation was related to the mesophyll cells. These results suggest the participation of the mesophyll in stomatal control. It is possible that stomatal opening in the intact leaf, to a greater or lesser extent. is dependent on an electrical signal or a chemical propagated from the mesophyll.
The effect of a number of factors on the opening of stomata in the intact leaf and in the isolated leaf epidermis of Commelina communis has been investigated. Stomata in the intact leaf opened wide in the light and closed rapidly on transfer to the dark. They were also sensitive to CO2. In contrast, stomata in isolated epidermis floated on an incubation solution containing 100 molm–3 KCl responded neither to light nor CO2. They opened as widely as those in the intact leaf when treated with fusicoccin.
Stomata in isolated epidermis opened almost as wide as those in the intact leaf when they were incubated with isolated mesophyll cells in the light. The solution in which the mesophyll cells were incubated was separated by centrifugation. The medium from cells previously incubated in the light caused the stomata in isolated epidermis to open but that from cells kept in the dark had no effect. A similar effect was observed when isolated chloroplasts were incubated with the isolated epidermis.However, the supernatant from the chloroplast suspension had no significant effect on stomatal opening.
These results indicate that the mesophyll plays an important role in stomatal opening in the light. The mesophyll appears to produce in the light, but not in the dark, a soluble compound which moves to the guard cells to bring about stomatal opening.The experiments with isolated chloroplasts suggest that this substance is a product of photosynthesis.
Competitive binding of antagonistic peptides fine-tunes stomatal patterning
by Lee J. S., Hnilova M., Maes M., Lin Y.-C. L., Putarjunan A., Han S.-K., Avila J.,Torii K. U. (2015)
in Nature 522,439–443(25 June 2015) doi:10.1038/nature14561
Figure 2: STOMAGEN overexpression on stomatal development in tmm hypocotyl epidermis with combinatorial loss-of-function in ER-family genes. – a–h, Representative confocal microscopy images of hypocotyl epidermis from 10-day-old light-grown transgenic Est::STOMAGEN (oestradiol-induced STOMAGEN) seedlings of tmm (a, b); tmm er (c, d); tmm erl1 erl2 (e, f); and tmm er erl1 erl2… – http://www.nature.com/nature/journal/v522/n7557/carousel/nature14561-f2.jpg
Abstract
During development, cells interpret complex and often conflicting signals to make optimal decisions. Plant stomata, the cellular interface between a plant and the atmosphere, develop according to positional cues, which include a family of secreted peptides called epidermal patterning factors (EPFs). How these signalling peptides orchestrate pattern formation at a molecular level remains unclear.
Here we report in Arabidopsis that Stomagen (also called EPF-LIKE9) peptide, which promotes stomatal development, requires ERECTA (ER)-family receptor kinases and interferes with the inhibition of stomatal development by the EPIDERMAL PATTERNING FACTOR 2 (EPF2)–ER module.
Both EPF2 and Stomagen directly bind to ER and its co-receptor TOO MANY MOUTHS. Stomagen peptide competitively replaced EPF2 binding to ER. Furthermore, application of EPF2, but not Stomagen, elicited rapid phosphorylation of downstream signalling components in vivo.
Our findings demonstrate how a plant receptor agonist and antagonist define inhibitory and inductive cues to fine-tune tissue patterning on the plant epidermis.
CHOR is required to restrict stomatal cell-lineage divisions. (A-C) Promoter activity of TMM(proTMM::GUS-GFP) in 6-day-old cotyledon epidermis of wild type (A) and chor (B,C). In chor,clusters of small cells (dots) show high GFP signals. Pavement cells in chor occasionally show incomplete cytokinesis (arrowhead). (D-F) Expression of MUTE (proMUTE::MUTE-GFP) in 6-day-old cotyledon epidermis of wild type (D) and chor (E, F). Clusters of small stomatal-lineage cells are in brackets. MUTE expression in adjacent meristemoids (asterisks) predicts the eventual formation of clustered stomata. (G-I) Expression of SPCH (proSPCH::SPCH-GFP) in 3-day-old cotyledon epidermis of wild type (G) and chor (H, I). More cells express GFP signals in chor than in wild type. (J-L) Effects of spch on the excessive stomatal-lineage divisions in chor. Shown is 10-day-old cotyledon epidermis of spch (J) and spch chor (K,L). Small, highly divided cells are no longer produced in spch chor (K,L), whereas incomplete cytokinesis of pavement cells is still evident (K,L, arrowheads). Cell peripheries were highlighted by propidium iodide (PI; A-F,J-L) or FM4-64 (G-I). A-F, G-I, and J-L were taken under the same magnification. Scale bars: 20 μm.
Dysregulation of cell-to-cell connectivity and stomatal patterning by loss-of-function mutation in Arabidopsis CHORUS (GLUCAN SYNTHASELIKE 8).
by Guseman J. M., Lee J. S., Bogenschutz N. L., Peterson K. M., Virata R. E., Xie B., Kanaoka M. M., Hong Z., Torii K. U. (2010)
in Development 137:1731–1741. – doi: 10.1242/dev.049197 –
Fig. 9. Model of phenotypical consequences of the loss of CHOR (GSL8) and signaling receptors. (A) In wild type, stomatal cell-fate determinants (cyan) are contained in a precursor cell, and inhibitory signals (red T bar) prevent its neighbors from adopting a stomatal-lineage fate. (B) In chor, cell-fate determinants (cyan) may leak through plasmodesmata, which results in excessive stomatal entry divisions. The functional inhibitory signals, however, are able to prevent the stomatal differentiation of neighbors at some level. This results in modest stomatal clusters. Incomplete cytokinesis also occurs in chor. (C) In the absence of both CHOR (GSL8) and signaling receptors (such as TMM), the loss (or reduction at the threshold level in er erl1) of inhibitory signals together with the leakage of cell-fate determinants leads to catastrophic stomatal patterning defects. – http://d2qiws50qrj9uc.cloudfront.net/content/develop/137/10/1731/F9.medium.gif
Summary
Patterning of stomata, valves on the plant epidermis, requires the orchestrated actions of signaling components and cell-fate determinants.
To understand the regulation of stomatal patterning, we performed a genetic screen using a background that partially lacks stomatal signaling receptors.
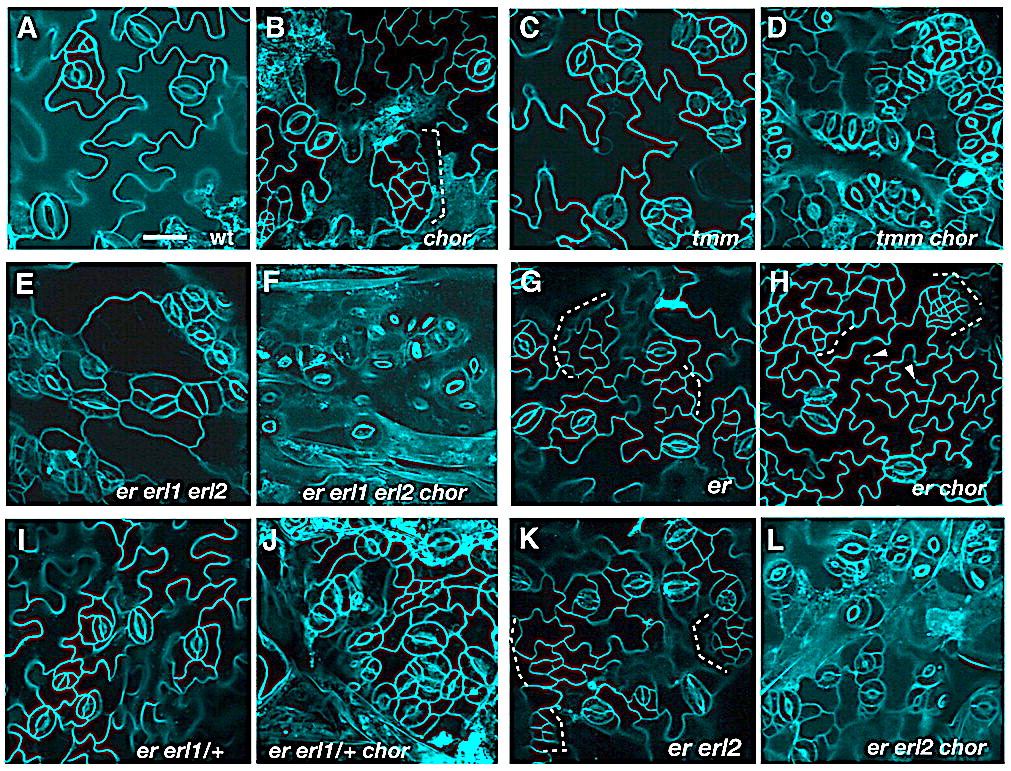
Genetic interactions of chor with stomatal patterning mutants. (A-L) Abaxial epidermis of rosette leaves (first pairs) from 12-day-old seedlings of the following genotypes: wild type (wt, A), chor (B), tmm (C), tmm chor (D), er erl1 erl2 (E), er erl1 erl2 chor (F), er (G), er chor (H), er erl1/+ (I), er erl1/+ chor (J), er erl2 (K) and er erl2 chor (L). chor in a wild-type background exhibits weak stomatal patterning defects and islands of small cells (dotted bracket; B), but the chormutation severely enhances stomatal clustering in tmm (C,D) and in the er erl1 erl2 triple mutant (E,F). er erl1/+ and er erl2 seedling leaves do not exhibit stomatal patterning defects (I,K); however, introduction of chor confers severe stomatal clustering defects (J,L). The er chordouble-mutant phenotype appeared additive, with increased small cells (dotted bracket) and incomplete cytokinesis (arrowheads, G,H). Images are taken under the same magnification. Scale bar: 20 μm.
Here, we report the isolation and characterization of chorus (chor), which confers excessive proliferation of stomatal-lineage cells mediated by SPEECHLESS (SPCH). chor breaks redundancy among three ERECTA family genes and strongly enhances stomatal patterning defects caused by loss-of-function in TOO MANY MOUTHS. chor seedlings also exhibit incomplete cytokinesis and growth defects, including disruptions in root tissue patterning and root hair cell morphogenesis.
CHOR encodes a putative callose synthase, GLUCAN SYNTHASE-LIKE 8 (GSL8), that is required for callose deposition at the cell plate, cell wall and plasmodesmata. Consistently, symplastic macromolecular diffusion between epidermal cells is significantly increased in chor, and proteins that do not normally move cell-to-cell, including a fluorescent protein-tagged SPCH, diffuse to neighboring cells.
Such a phenotype is not a general trait caused by cytokinesis defects.
Our findings suggest that the restriction of symplastic movement might be an essential step for the proper segregation of cell-fate determinants during stomatal development.
During development, cells interpret complex and often conflicting signals to make optimal decisions. Plant stomata, the cellular interface between a plant and the atmosphere, develop according to positional cues, which include a family of secreted peptides called epidermal patterning factors (EPFs). How these signalling peptides orchestrate pattern formation at a molecular level remains unclear.
Here we report in Arabidopsis that Stomagen (also called EPF-LIKE9) peptide, which promotes stomatal development, requires ERECTA (ER)-family receptor kinases and interferes with the inhibition of stomatal development by the EPIDERMAL PATTERNING FACTOR 2 (EPF2)–ER module. Both EPF2 and Stomagen directly bind to ER and its co-receptor TOO MANY MOUTHS. Stomagen peptide competitively replaced EPF2 binding to ER. Furthermore, application of EPF2, but not Stomagen, elicited rapid phosphorylation of downstream signalling components in vivo.
Our findings demonstrate how a plant receptor agonist and antagonist define inhibitory and inductive cues to fine-tune tissue patterning on the plant epidermis.
In the age of tablet computers and smart phones, it’s easy to feel inundated and overloaded by information. But on a cellular level, this bombardment is business as usual, and a team of University of Washington researchers has identified a mechanism that some plant cells use to receive complex and contradictory messages from their neighbors.
As they report in a paper published online June 17 in Nature, the team led by UW biology professor and senior author Keiko Torii made its discovery as they explored how plants organize cellular structures on their surface.
Like other multicellular creatures, plants must coordinate activity among many different types of cells and tissues. Messages, demands, warnings and alerts shuttle among cells near and far. These messages determine what jobs cells take on and how they work together to build and maintain tissues and organs. As plants grow, they also use this information to decide where new structures like leaves or roots should go.
Torii, lead author Jin Suk Lee and their colleagues focused on how plants decide where to place stomata: tiny, two-cell openings on the surface that connect the plant’s interior with the outside world. Critical for water and gas exchange, stomata develop on the plant’s surface based largely on signals they receive from neighboring cells.
Plants must grow and distribute their stomata evenly on the surface because too many or too few can disrupt water balance or photosynthesis.
Plants exposed to excess amounts of Stomagen – the pro-stomata signal – develop extra stomata in clusters.Jin Suk Lee, Nature
Lee and Torii studied two signals that plant cells release to control where stomata go. These signals are actually proteins, or small molecules that help cells do work and communicate with one another. One is called Stomagen, which promotes stomata development. The other protein messenger — known by its acronym EPF2 — opposes Stomagen by preventing stomata formation.








{kind=link}
{kind=link}
You must be logged in to post a comment.