The science of the stomata of plants: a continuously growing list of references, abstracts and illustrations, helping researchers to data on publications.
Attention is drawn to the close similarity in epidermal structure between Austrobaileya (Austrobaileyaceae) and Schisandraceae and to the presence in both families of the laterocytic stomatal type.
As carbon dioxide levels have risen during the last 150 years, the density of pores that allow plants to breathe has dwindled by 34 percent, restricting the amount of water vapor the plants release to the atmosphere, report scientists from Indiana University Bloomington and Utrecht University in the Netherlands in an upcoming issue of the Proceedings of the National Academy of Sciences (now online).
In a separate paper, also to be published by PNAS, many of the same scientists describe a model they devised that predicts doubling today’s carbon dioxide levels will dramatically reduce the amount of water released by plants.
The scientists gathered their data from a diversity of plant species in Florida, including living individuals as well as samples extracted from herbarium collections and peat formations 100 to 150 years old.
“The increase in carbon dioxide by about 100 parts per million has had a profound effect on the number of stomata and, to a lesser extent, the size of the stomata,” said Research Scientist in Biology and Professor Emeritus in Geology David Dilcher, the two papers’ sole American coauthor. “Our analysis of that structural change shows there’s been a huge reduction in the release of water to the atmosphere.”
Most plants use a pore-like structure called stomata (singular: stoma) on the undersides of leaves to absorb carbon dioxide from the air. The carbon dioxide is used to build sugars, which can be used by the plant as energy or for incorporation into the plants’ fibrous cell walls. Stomata also allow plants to “transpire” water, or release water to the atmosphere. Transpiration helps drive the absorption of water at the roots, and also cools the plants in the same way sweating cools mammals.
If there are fewer stomata, or the stomata are closed more of the day, gas exchange will be limited — transpiration included.
Rising carbon dioxide is causing plants to have fewer pores, releasing less water to the atmosphere
Date:March 4, 2011
Source:Indiana University
Summary:
As carbon dioxide levels have risen during the last 150 years, the density of pores that allow plants to breathe has dwindled by 34 percent, restricting the amount of water vapor the plants release to the atmosphere, report scientists.
As carbon dioxide levels have risen during the last 150 years, the density of pores that allow plants to breathe has dwindled by 34 percent, restricting the amount of water vapor the plants release to the atmosphere, report scientists from Indiana University Bloomington and Utrecht University in the Netherlands in an upcoming issue of the Proceedings of the National Academy of Sciences.
Journal References:
E. I. Lammertsma, H. J. de Boer, S. C. Dekker, D. L. Dilcher, A. F. Lotter, F. Wagner-Cremer. Global CO2 rise leads to reduced maximum stomatal conductance in Florida vegetation. Proceedings of the National Academy of Sciences, 2011; DOI: 10.1073/pnas.1100371108
H. J. de Boer, E. I. Lammertsma, F. Wagner-Cremer, D. L. Dilcher, M. J. Wassen, S. C. Dekker. Climate forcing due to optimization of maximal leaf conductance in subtropical vegetation under rising CO2. Proceedings of the National Academy of Sciences, 2011; DOI: 10.1073/pnas.1100555108
In grapevine, the penetration and sporulation of Plasmopara viticola occur via stomata, suggesting functional relationships between guard cells and the pathogen. This assumption was supported by our first observation that grapevine (Vitis vinifera cv. Marselan) cuttings infected by P. viticola wilted more rapidly than healthy ones when submitted to water starvation.
Here, complementary approaches measuring stomatal conductance and infrared thermographic and microscopic observations were used to investigate stomatal opening/closure in response to infection.
In infected leaves, stomata remained open in darkness and during water stress, leading to increased transpiration. This deregulation was restricted to the colonized area, was not systemic and occurred before the appearance of symptoms.
Cytological observations indicated that stomatal lock-open was not related to mechanical forces resulting from the presence of the pathogen in the substomatal cavity.
In contrast to healthy leaves, stomatal closure in excised infected leaves could not be induced by a water deficit or abscisic acid (ABA) treatment. However, ABA induced stomatal closure in epidermal peels from infected leaves, indicating that guard cells remained functional.
These data indicate that the oomycete deregulates guard cell functioning, causing significant water losses. This effect could be attributed to a non-systemic compound, produced by the oomycete or by the infected plant, which inhibits stomatal closure or induces stomatal opening; or a reduction of the back-pressure exerted by surrounding epidermal cells.
Stomata, natural pores bordered by guard cells, regulate transpiration and gas exchanges between plant leaves and the atmosphere. These natural openings also constitute a way of penetration for microorganisms.
In plants, the perception of potentially pathogenic microorganisms or elicitors of defense reactions induces a cascade of events, including H2O2 production, that allows the activation of defense genes, leading to defense reactions.
Similar signaling events occur in guard cells in response to the perception of abscisic acid (ABA), leading to stomatal closure.
Moreover, few elicitors were reported to induce stomatal closure in Arabidopsis and Vicia faba leaves. Because responses to ABA and elicitors share common signaling events, it led us to question whether stomatal movements and H2O2 production in guard cells could play a key role in elicitor-induced protection against pathogens that use stomata for infection.
This study was performed using the grapevine– Plasmopara viticola pathosystem. Using epidermal peels, we showed that, as for ABA, the elicitor-induced stomatal closure is mediated by reactive oxygen species (ROS) production in guard cells.
In plants, we observed that the protection against downy mildew induced by some elicitors is probably not due only to effects on stomatal movements or to a guard-cell-specific activation of ROS production.
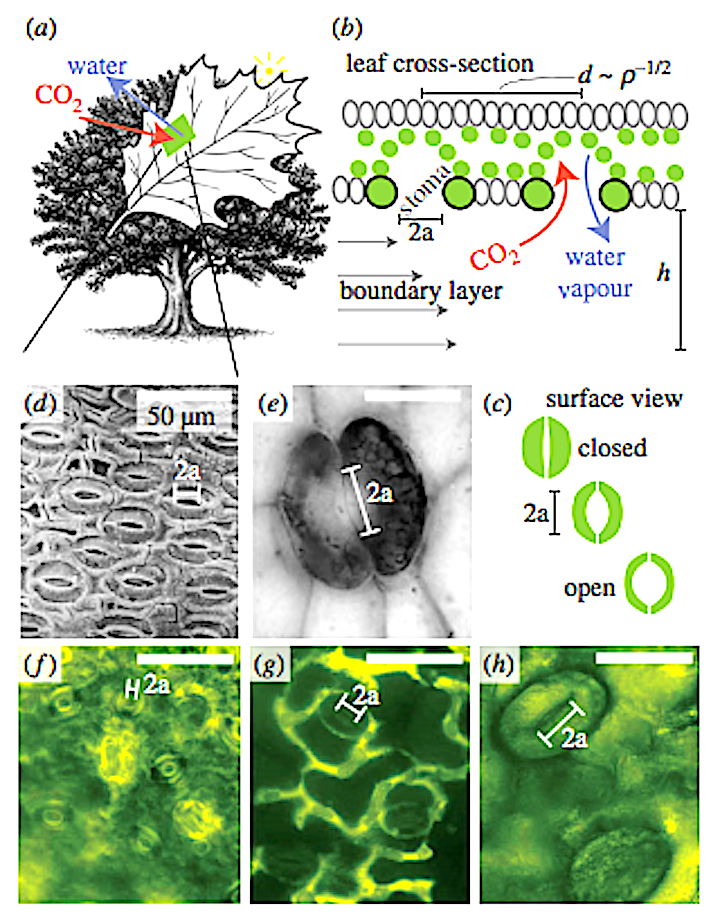
Figure 1. Gas exchange for photosynthesis and respiration takes place by way of small stoma pores on the surface of plant leaves. (a,b) Stoma apertures are approximately circular with radius a, and a boundary layer of thickness h separates the pores from the bulk atmosphere. The pore-to-pore distance is d ¼ r21/2, where r is the stomatal density. (c) The action of guard cells allows for opening and closing of the pore in response to environmental cues. When fully open pores are often circular in shape with radius approximately one-sixth of the total stoma complex length. Panels (d–h) illustrate the diversity of stoma size and density. (d) Swillingtonia denticulata (image modified from [1]), (e) Aglaophyton major (image modified from [2]) ( f ) Citrus reticulata, (g) Gossypium hirsutum and (h) Nephrolepis exaltata.
Stomatal design principles in synthetic and real leaves
by Zwieniecki M., Haaning K. S., Boyce C. K., Jensen K. H. (2016)
Maciej A. Zwieniecki 1 , Katrine S. Haaning 2 , C. Kevin Boyce 3 , Kaare H. Jensen 2
1 Department of Plant Sciences, University of California, Davis, CA 95616, USA
2 Department of Physics, Technical University of Denmark, DK-2800 Kgs. Lyngby, Denmark
3 Department of Geological Sciences, Stanford University, Stanford, CA 94305, USA
Stomata are portals in plant leaves that control gas exchange for photosynthesis, a process fundamental to life on Earth. Gas fluxes and plant productivity depend on external factors such as light, water and CO2 availability and on the geometrical properties of the stoma pores.
The link between stoma geometry and environmental factors has informed a wide range of scientific fields—from agriculture to climate science, where observed variations in stoma size and density are used to infer prehistoric atmospheric CO2 content.
However, the physical mechanisms and design principles responsible for major trends in stomatal patterning are not well understood. Here, we use a combination of biomimetic experiments and theory to rationalize the observed changes in stoma geometry.
We show that the observed correlations between stoma size and density are consistent with the hypothesis that plants favour efficient use of space and maximum control of dynamic gas conductivity, and that the capacity for gas exchange in plants has remained constant over at least the last 325 Myr.
Our analysis provides a new measure to gauge the relative performance of species based on their stomatal characteristics.
Figs. I-S.-Representative standards for scoring of the degree of potassiun staining in guard cells: I, 5%; 2, 10%; 3, 20%; 4, 35%; 5, 50%; 6, 60%; 7, SO%; and S, 100%. In Figure 7 the unstained region is that occupied by the guard cell nucleus; generally, however, the intra. cellular location of the stain is not a good indicator of the original location of the potassiun (Macallum 1905).
Aspects of potassium accumulation by stomata of Vicia faba.
Photo credit: CSIRO -Figs. 9-12.-Epidermal strips stained for potassium (see text) to show potassium in intact epidermal cells (black precipitate within the cell boundary): 9, low-power view showing a patch of intact epidermal cells; 10, 11. and 12, respectively, are high.power views of stomata surrounded entirely, on one side, and not at all by intact epidermal cells.
Abstract
Potassium accumulation by guard cells during the light opening of Vicia fabastomata in epidermal strips was determined quantitatively with 42K and 86Rb tracers. The sodium cobaltinitrite stain for potassium was also used. Particular attention was paid to errors arising from the presence of intact epidermal cells.
When epidermal strips obtained from darkened leaves were floated on potassium chloride solution (usually 10 mM) in light plus C02-free air, guard cell potassium content and stomatal aperture increased in parallel, both approaching a maximum after about 300 min.
Flux measurement suggested that the steady maximum potassium content (and aperture) arises because of a decline in the initially high potassium influx.
With opening of stomata in the light, external calcium reduced in a parallel manner both opening and potassium uptake at potassium concentrations ranging from nearly zero to 50 mM. Also, maximum stomatal opening and potassium accumulation in the presence or absence of calcium bore linear relationships to log external potassium concentration.
These results are discussed in relation to potassium accumulation in other plant systems. In experiments with epidermal strips, guard cell potassium content showed a consistent linear relationship to stomatal aperture.
For 16 experiments spanning two years and many treatments affecting aperture, the mean slope of this relationship was 2·6 nmoles cm-2 p.m-l. This is equivalent to a change in potassium concentration of 40 mM p.m-l. The increase in guard cell osmotic pressure attributable to potassium plus anion accumulation is estimated to be 1·5 bars p.m-l, which is a major portion of the observed increase in osmotic pressure with opening (2, 0 bars p.m-l).
Stomatal characteristics of an extinct Cretaceous conifer, Pseudofrenelopsis parceramosa (Fontaine) Watson, are used to reconstruct atmospheric carbon dioxide (pCO2) over a time previously inferred to exhibit major fluctuations in this greenhouse gas.
Samples are from non-marine to marine strata of the Wealden and Lower Greensand Groups of England and the Potomac Group of the eastern United States, of Hauterivian to Albian age (136–100 Ma).
Atmospheric pCO2 is estimated from the ratios between stomatal indices of fossil cuticles and those from four modern analogs (nearest living equivalent plants). Using this approach, and two calibration methods to explore ranges, results show relatively low and only slightly varying pCO2 over the Hauterivian–Albian interval: a low of ∼560–960 ppm in the early Barremian and a high of ∼620–1200 ppm in the Albian.
Data from the Barremian Wealden Group yield pCO2 values indistinguishable from a soil-carbonate–based estimate from the same beds. The new pCO2 estimates are compatible with sedimentological and oxygen-isotope evidence for relatively cool mid-Cretaceous climates.
Development of stomata, valves on the plant epidermis for optimal gas exchange and water control, is fine-tuned by multiple signaling peptides with unique, overlapping, or antagonistic activities.
EPIDERMAL PATTERNING FACTOR1 (EPF1) is a founding member of the secreted peptide ligands enforcing stomatal patterning. Yet, its exact role remains unclear.
Here, we report that EPF1 and its primary receptor ERECTA-LIKE1 (ERL1) target MUTE, a transcription factor specifying the proliferation-to-differentiation switch within the stomatal cell lineages.
In turn, MUTE directly induces ERL1.
The absolute co-expression of ERL1 and MUTE, with the co-presence of EPF1, triggers autocrine inhibition of stomatal fate.
During normal stomatal development, this autocrine inhibition prevents extra symmetric divisions of stomatal precursors likely owing to excessive MUTE activity.
Our study reveals the unexpected role of self-inhibition as a mechanism for ensuring proper stomatal development and suggests an intricate signal buffering mechanism underlying plant tissue patterning.
This experiment compares stomata density of the epiphytic Pleurothallis aristata and Maxillaria sp. orchids under experimental conditions of dry and humid environments.
Pleurothallis aristata is in the sub-tribe Pleurothallidinae and lacks pseudobulbs, while Maxillaria sp. has pseudobulbs.
The study seeks to determine what differences in stomata density exist between the two species, and if there is a difference in mean percent stomata open in humid and dry environments.
The study takes stomata impressions from the leaves of twenty individuals of each species using clear nail polish. The results show a significant difference in stomata density between the Pleurothallis aristata and the Maxillaria sp. (Rank Sum Test: t=55, n1=10, n2=10, p<0.05).
Additionally, both species have a higher percentage of open stomata in humid environments than in dry environments (Wilcoxon sign rank test). An explanation for these results is that Maxillaria sp. has a pseudobulb for water storage, has a larger leaf surface area, and therefore has higher stomata density.
The study showed both species closed their stomata in drier conditions in order to reduce water loss and desiccation.
The results of this experiment help demonstrate how different orchid species function in humid and dry environments, and their ability to succeed in the event of global climate change and shifting of biomes.










{kind=link}
You must be logged in to post a comment.