The science of the stomata of plants: a continuously growing list of references, abstracts and illustrations, helping researchers to data on publications.
It has been successfully demonstrated, using epidermis explants of sugar beet (Beta vulgaris L.), that stomatal guard cells retain full totipotent capacity. Despite having one of the highest degrees of morphological adaptation and a unique physiological specialization, it is possible to induce a re-expression of full (embryogenic) genetic potential in these cells in situ by reversing their highly differentiated nature to produce regenerated plants via a callus stage.
The importance of these findings both to stomatal research and to our understanding of cytodifferentiation in plants is discussed.
Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor: Partitioning stomatal responses to drought.
by Rodriguez-Dominguez C. M.,
Buckley T. N.,
Egea G.,
Diaz-Espejo A.
(2016)
in Plant Cell and Environment · June 2016 – DOI: 10.1111/pce.12774
Reduced stomatal conductance (gs) during soil drought in angiosperms may result from effects of leaf turgor on stomata, and/or factors that do not directly depend on leaf turgor, including root-derived abscisic acid (ABA) signals.
To quantify the roles of leaf-turgor-mediated and leaf-turgor-independent mechanisms in gs decline during drought, we measured drought responses of gs and water relations in three woody species (almond, grapevine and olive) under a range of conditions designed to generate independent variation in leaf and root turgor, including diurnal variation in evaporative demand and changes in plant hydraulic conductance and leaf osmotic pressure.
We then applied these data to a process-based gs model and used a novel method to partition observed declines in gs during drought into contributions from each parameter in the model. Soil drought reduced gs by 63-84% across species, and the model reproduced these changes well (r2 = 0.91, p < 0.0001, n = 44) despite having only a single fitted parameter.
Our analysis concluded that responses mediated by leaf turgor could explain over 87% of the observed decline in gs across species, adding to a growing body of evidence that challenges the root-ABA-centric model of stomatal responses to drought.
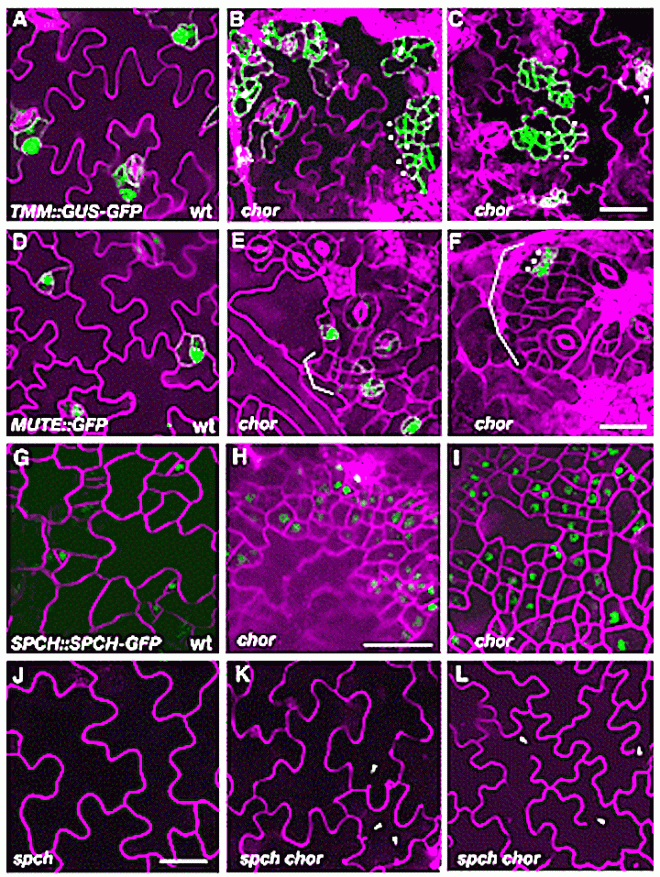
CHOR is required to restrict stomatal cell-lineage divisions. (A-C) Promoter activity of TMM(proTMM::GUS-GFP) in 6-day-old cotyledon epidermis of wild type (A) and chor (B,C). In chor,clusters of small cells (dots) show high GFP signals. Pavement cells in chor occasionally show incomplete cytokinesis (arrowhead). (D-F) Expression of MUTE (proMUTE::MUTE-GFP) in 6-day-old cotyledon epidermis of wild type (D) and chor (E, F). Clusters of small stomatal-lineage cells are in brackets. MUTE expression in adjacent meristemoids (asterisks) predicts the eventual formation of clustered stomata. (G-I) Expression of SPCH (proSPCH::SPCH-GFP) in 3-day-old cotyledon epidermis of wild type (G) and chor (H, I). More cells express GFP signals in chor than in wild type. (J-L) Effects of spch on the excessive stomatal-lineage divisions in chor. Shown is 10-day-old cotyledon epidermis of spch (J) and spch chor (K,L). Small, highly divided cells are no longer produced in spch chor (K,L), whereas incomplete cytokinesis of pavement cells is still evident (K,L, arrowheads). Cell peripheries were highlighted by propidium iodide (PI; A-F,J-L) or FM4-64 (G-I). A-F, G-I, and J-L were taken under the same magnification. Scale bars: 20 μm.
Dysregulation of cell-to-cell connectivity and stomatal patterning by loss-of-function mutation in Arabidopsis CHORUS (GLUCAN SYNTHASELIKE 8).
by Guseman J. M., Lee J. S., Bogenschutz N. L., Peterson K. M., Virata R. E., Xie B., Kanaoka M. M., Hong Z., Torii K. U. (2010)
in Development 137:1731–1741. – doi: 10.1242/dev.049197 –
Fig. 9. Model of phenotypical consequences of the loss of CHOR (GSL8) and signaling receptors. (A) In wild type, stomatal cell-fate determinants (cyan) are contained in a precursor cell, and inhibitory signals (red T bar) prevent its neighbors from adopting a stomatal-lineage fate. (B) In chor, cell-fate determinants (cyan) may leak through plasmodesmata, which results in excessive stomatal entry divisions. The functional inhibitory signals, however, are able to prevent the stomatal differentiation of neighbors at some level. This results in modest stomatal clusters. Incomplete cytokinesis also occurs in chor. (C) In the absence of both CHOR (GSL8) and signaling receptors (such as TMM), the loss (or reduction at the threshold level in er erl1) of inhibitory signals together with the leakage of cell-fate determinants leads to catastrophic stomatal patterning defects. – http://d2qiws50qrj9uc.cloudfront.net/content/develop/137/10/1731/F9.medium.gif
Summary
Patterning of stomata, valves on the plant epidermis, requires the orchestrated actions of signaling components and cell-fate determinants.
To understand the regulation of stomatal patterning, we performed a genetic screen using a background that partially lacks stomatal signaling receptors.
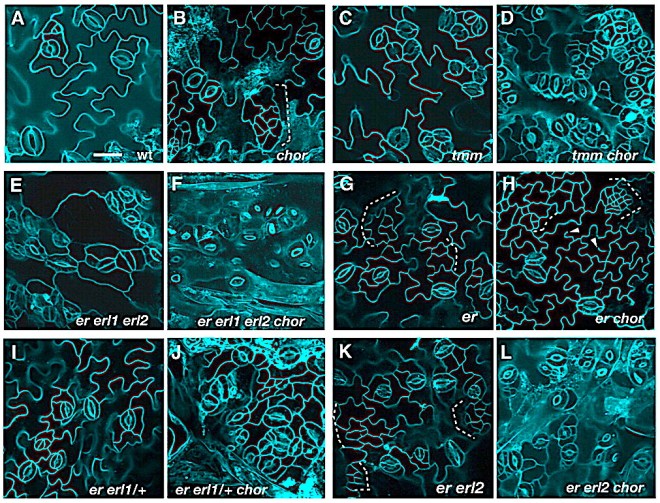
Genetic interactions of chor with stomatal patterning mutants. (A-L) Abaxial epidermis of rosette leaves (first pairs) from 12-day-old seedlings of the following genotypes: wild type (wt, A), chor (B), tmm (C), tmm chor (D), er erl1 erl2 (E), er erl1 erl2 chor (F), er (G), er chor (H), er erl1/+ (I), er erl1/+ chor (J), er erl2 (K) and er erl2 chor (L). chor in a wild-type background exhibits weak stomatal patterning defects and islands of small cells (dotted bracket; B), but the chormutation severely enhances stomatal clustering in tmm (C,D) and in the er erl1 erl2 triple mutant (E,F). er erl1/+ and er erl2 seedling leaves do not exhibit stomatal patterning defects (I,K); however, introduction of chor confers severe stomatal clustering defects (J,L). The er chordouble-mutant phenotype appeared additive, with increased small cells (dotted bracket) and incomplete cytokinesis (arrowheads, G,H). Images are taken under the same magnification. Scale bar: 20 μm.
Here, we report the isolation and characterization of chorus (chor), which confers excessive proliferation of stomatal-lineage cells mediated by SPEECHLESS (SPCH). chor breaks redundancy among three ERECTA family genes and strongly enhances stomatal patterning defects caused by loss-of-function in TOO MANY MOUTHS. chor seedlings also exhibit incomplete cytokinesis and growth defects, including disruptions in root tissue patterning and root hair cell morphogenesis.
CHOR encodes a putative callose synthase, GLUCAN SYNTHASE-LIKE 8 (GSL8), that is required for callose deposition at the cell plate, cell wall and plasmodesmata. Consistently, symplastic macromolecular diffusion between epidermal cells is significantly increased in chor, and proteins that do not normally move cell-to-cell, including a fluorescent protein-tagged SPCH, diffuse to neighboring cells.
Such a phenotype is not a general trait caused by cytokinesis defects.
Our findings suggest that the restriction of symplastic movement might be an essential step for the proper segregation of cell-fate determinants during stomatal development.
Pathogen-induced stomatal closure is part of the plant innate immune response. Phytopathogens using stomata as a way of entry into the leaf must avoid the stomatal response of the host.
In this article, we describe a factor secreted by the bacterial phytopathogen Xanthomonas campestris pv campestris (Xcc) capable of interfering with stomatal closure induced by bacteria or abscisic acid (ABA).
We found that living Xcc, as well as ethyl acetate extracts from Xcc culture supernatants, are capable of reverting stomatal closure induced by bacteria, lipopolysaccharide, or ABA. Xcc ethyl acetate extracts also complemented the infectivity of Pseudomonas syringae pv tomato (Pst) mutants deficient in the production of the coronatine toxin, which is required to overcome stomatal defense.
By contrast, the rpfF and rpfC mutant strains of Xcc, which are unable to respectively synthesize or perceive a diffusible molecule involved in bacterial cell-to-cell signaling, were incapable of reverting stomatal closure, indicating that suppression of stomatal response by Xcc requires an intact rpf/diffusible signal factor system.
In addition, we found that guard cell-specific Arabidopsis (Arabidopsis thaliana) Mitogen-Activated Protein Kinase3 (MPK3) antisense mutants were unresponsive to bacteria or lipopolysaccharide in promotion of stomatal closure, and also more sensitive to Pstcoronatine-deficient mutants, showing that MPK3 is required for stomatal immune response.
Additionally, we found that, unlike in wild-type Arabidopsis, ABA-induced stomatal closure in MPK3 antisense mutants is not affected by Xcc or by extracts from Xccculture supernatants, suggesting that the Xcc factor might target some signaling component in the same pathway as MPK3.
Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor
by Rodriguez-Dominguez C. M.,
Buckley T. N.,
Egea G.,
de Cires A.,
Hernandez-Santana V.,
Martorell S.,
Diaz-Espejo A.
(2016) –
in Wiley Online LIbrary: Browse Accepted Articles
Accepted, unedited articles published online and citable. The final edited and typeset version of record will appear in future.
DOI: 10.1111/pce.12774
Abstract
Reduced stomatal conductance (gs) during soil drought in angiosperms may result from effects of leaf turgor on stomata, and/or factors that do not directly depend on leaf turgor, including root-derived abscisic acid (ABA) signals.
To quantify the roles of leaf-turgor-mediated and leaf-turgor-independent mechanisms in gs decline during drought, we measured drought responses of gs and water relations in three woody species (almond, grapevine and olive) under a range of conditions designed to generate independent variation in leaf and root turgor, including diurnal variation in evaporative demand and changes in plant hydraulic conductance and leaf osmotic pressure.
We then applied these data to a process-based gs model and used a novel method to partition observed declines in gs during drought into contributions from each parameter in the model.
Soil drought reduced gs by 63-84% across species, and the model reproduced these changes well (r2 = 0.91, p < 0.0001, n = 44) despite having only a single fitted parameter.
Our analysis concluded that responses mediated by leaf turgor could explain over 87% of the observed decline in gs across species, adding to a growing body of evidence that challenges the root-ABA-centric model of stomatal responses to drought.
• MAP kinases have been linked to guard cell signalling. Arabidopsis thaliana MAP Kinase 3 (MPK3) is known to be activated by abscisic acid (ABA) and hydrogen peroxide (H2O2), which also control stomatal movements.
• We therefore studied the possible role of MPK3 in guard cell signalling through guard cell-specific antisense inhibition of MPK3 expression.
• Such transgenic plants contained reduced levels of MPK3 mRNA in the guard cells and displayed partial insensitivity to ABA in inhibition of stomatal opening, but responded normally to this hormone in stomatal closure. However, ABA-induced stomatal closure was reduced compared with controls when cytoplasmic alkalinization was prevented with sodium butyrate. MPK3 antisense plants were less sensitive to exogenous H2O2, both in inhibition of stomatal opening and in promotion of stomatal closure, thus MPK3 is required for the signalling of this compound. ABA-induced H2O2 synthesis was normal in these plants, indicating that MPK3 probably acts in signalling downstream of H2O2.
• These results provide clear evidence for the important role of MPK3 in the perception of ABA and H2O2 in guard cells.
• Morphological and physiological attributes were assessed to elucidate the underlying mechanisms of ozone (O3) sensitivity in a highly sensitive species, cutleaf coneflower (Rudbeckia laciniata var. digitata).
• Foliage at the same height in the canopy on paired O3-sensitive and -insensitive cutleaf coneflowers was assessed for level of foliar symptoms, stomatal density,stomatal responsiveness to dynamic changes in light and leaf-to-air vapor pressure deficit (VPD), steady-state responses to light and CO2, intrinsic transpirational efficiency, and plant water balance.
• There were no morphological differences between the sensitivity types that might have contributed to greater O3uptake in sensitive individuals. Stomata of sensitive plants were less responsive than those of insensitive plants to experimentally increased and decreased light intensities, and to increased VPD. O3-insensitive plants had greater intrinsic transpirational efficiencies, greater maximum assimilation rates under saturating CO2 and light, and greater carboxylation rates.
• Different physiological attributes vary independently within an individual plant, which collectively confer sensitivity or insensitivity to O3 injury.




















{kind=link}
You must be logged in to post a comment.